Une étude sur la sentience des insectes, publiée le 14 novembre 2022, conclut que certains insectes ressentiraient la douleur.

Une revue systématique de la littérature scientifique publiée le 14 novembre 2022 conclut en faveur de l’idée générale que les insectes (ou au moins certains d’entre eux) ressentiraient la douleur. Deux des auteurs de cette revue sont d’ailleurs co-auteurs du récent rapport de la London School of Economics, commissionné par le gouvernement britannique pour évaluer la sensibilité à la douleur chez les crustacés et les céphalopodes. Retour sur cette nouvelle publication et plus largement, sur la sentience des insectes et sur les méthodes d’étude de la sentience des animaux.
Lire aussi : Le mot sentience entre dans le Larousse 2020 (n° 102)
I. Introduction: la notion de sentience et rappels éthiques
a) La sentience
La sentience peut se définir comme la capacité à ressentir (Gibbons et al., 2022 ; Birch et al., 2021). Elle se distingue de la simple conscience dite « phénoménale » – la capacité à avoir des états mentaux subjectifs comme par exemple le ressenti perceptif associé à la vision des couleurs. La sentience renvoie en effet plus spécifiquement à la capacité à ressentir des états mentaux colorés par une valence positive (ressentis agréables comme le plaisir, la joie, la bonne humeur, la satiété etc.) ou négative (ressentis désagréables comme la douleur, la peur, l’anxiété, ou la faim). Dans sa forme minimale, la sentience renvoie essentiellement à la capacité à ressentir la douleur et le plaisir.
Le mot français « sensible » est souvent employé comme une traduction du terme anglais « sentience » mais n’en est pas un synonyme parfait. En effet, la sentience implique cette dimension de ressenti subjectif conscient positif ou négatif, tandis que la sensibilité peut se limiter à la capacité à détecter des signaux. Cela permet de dire qu’une plante et une pellicule photographique sont des êtres sensibles (à la lumière), sans pour autant être conscientes ni sentientes, comme l’explique le neurobiologiste Georges Chapouthier.
Si la sensibilité à la douleur est un critère suffisant pour qu’un individu soit considéré comme sentient, ce n’est pas obligatoirement un critère nécessaire*. De fait, la sentience ne se limite pas à cette seule dimension. La capacité à éprouver subjectivement des sensations agréables comme le plaisir et la satiété en est un autre aspect très important.
Dans ses formes plus élaborées, entres d’autres formes d’états mentaux subjectifs en plus des ressentis sensoriels, la sentience renvoie aussi à la capacité à ressentir des émotions positives ou négatives. Un continuum de complexité existe dans ce domaine chez les différents animaux (non humains et humains). Cela va du relativement basique comme la peur, la joie, le dégoût, l’attachement et la tristesse, au très complexe comme la honte, la nostalgie, ou le sentiment d’accomplissement de soi.
* En pratique toutefois, des cas d’animaux capables de ressentir des états mentaux positifs mais incapables de ressentir la douleur (sauf pathologie particulière) n’ont jamais été décrits, à la connaissance de l’auteur.
b) Implications éthiques et perspectives situées des chercheurs
La sentience d’un groupe d’individus a des implications éthiques majeures. En effet, elle leur confère la possibilité d’avoir des intérêts : leur vie peut se passer bien ou mal. La manière dont elle est étudiée et débattue sur le plan scientifique révèle souvent de manière implicite certains partis pris subjectifs. Les perspectives toujours « situées »* des différents auteurs étant implicitement marquées par des visions philosophiques ou politiques.
*La notion de connaissance située renvoie à l’idée que le contexte sociétal dans lequel sont produites les connaissances, et notamment la position sociale des chercheurs et leurs opinions politiques (leur « perspective située »), exerce nécessairement une influence plus ou moins marquée de manière implicite sur les savoirs produits.
Cela se voit tout particulièrement quant aux hypothèses par défaut retenues, et dans la manière de gérer les incertitudes. En l’absence d’étude sur la sentience d’un groupe d’animaux, l’hypothèse par défaut la plus précautionneuse est-elle de présumer la sentience ou l’insensibilité ? En cas d’incertitude sur les conclusions, doit-on ou ne doit-on pas donner le bénéfice du doute au groupe étudié ? Devrait-on alors le traiter comme si ses membres étaient sentients ? Doit-on raisonner uniquement en recherchant chez les animaux non-humains les mêmes structures biologiques impliquées dans la sentience chez les humains ? Ou peut-on considérer que l’absence de ces structures n’est pas une preuve d’insensibilité, car des structures biologiquement différentes des nôtres pourraient avoir des similarités fonctionnelles ? Cela permettrait-il alors d’aboutir à un ressenti similaire au nôtre par des processus différents des nôtres ? Et enfin, à quel niveau de protection voire à quels droits la sentience devrait-elle permettre d’accéder ? Le caractère vulnérable des individus (humains ou non humains) d’un groupe, leur asservissement à des pratiques d’exploitation lucratives, l’existence de discours idéologiques sur leur infériorité morale forment un contexte sociétal qui rend impossible une réelle neutralité de point de vue.
Par exemple, au début du XXe siècle, certains groupes humains, notamment les très jeunes enfants et les personnes non-verbales en situation de handicap mental, étaient jugés par une large partie de la communauté scientifique comme insensibles à la douleur, et traités comme tels par le personnel médical. Aujourd’hui encore, la prise en charge de la douleur de ces humains vulnérables n’est pas systématique au niveau mondial.
Lire aussi : Vers une définition plus inclusive de la douleur (n° 107)
c) Les avancées précédentes sur la sentience animale
Au XVIIe siècle, René Descartes, avec sa fameuse thèse de l’animal-machine, considérait les animaux non-humains comme dépourvus de tout ressenti subjectif (des débats existent sur l’interprétation de la pensée de Descartes sur ce sujet). Dans les années 1960 avaient lieu les premiers débats modernes sur la sentience et le bien-être des animaux d’élevage en Angleterre. Au début des années 2000 étaient publiés les tous premiers travaux sur la sensibilité à la douleur chez les poissons. Au départ assez controversée, elle est aujourd’hui bien établie. En 2012, un groupe de scientifiques proclamait la Déclaration de Cambridge sur la conscience, visibilisant un certain consensus quant au fait qu’un grand nombre d’animaux (notamment les mammifères, les oiseaux et les pieuvres) possèdent les substrats neurologiques de la conscience. Par la suite, en 2017, un rapport d’expertise collective de l’Inrae concluait lui aussi que les animaux vertébrés sont doués d’un certain niveau de conscience.
Les considérations à propos des invertébrés sont encore plus récentes. Ainsi, l’année dernière seulement, le gouvernement britannique intégrait les céphalopodes et les crustacés décapodes dans le champ d’application de certains textes de protection animale (la « Sentience bill »). Cela fit suite à la publication d’un rapport d’expertise de la London School of Economics concluant qu’ils sont sensibles à la douleur.
Maintenant qu’un relatif consensus a été atteint concernant les crustacés décapodes et les céphalopodes, c’est aujourd’hui les insectes qui constituent le nouveau taxon au cœur des travaux scientifiques sur la sentience. Le groupe des insectes est d’ailleurs l’une des classes de l’embranchement des arthropodes auquel appartiennent aussi les crustacés et les arachnides (araignées, scorpions). Il en existe 1,3 million d’espèces décrites, mais on estime entre 5 et 80 millions le nombre d’espèces total, représentant 86 % de la biodiversité animale en nombre d’espèces.
II – Les insectes sont-ils conscients ?
De plus en plus, la conscience est pensée comme un phénomène protéiforme, recouvrant un continuum de niveaux de conscience divers. La conscience « phénoménale » en est le niveau le plus basique, renvoyant simplement à la capacité à avoir des états mentaux subjectifs, indépendamment du contenu et de la complexité de ces états mentaux.
Lire aussi : Qu’est-ce que la conscience ? (n° 94)
La Déclaration de Cambridge de 2012 réfutait l’idée selon laquelle le néocortex* serait un prérequis de la conscience, ouvrant la porte à la possibilité que des animaux autres que les mammifères, y compris certains invertébrés, puissent être dotés de conscience. Pour autant, ce texte ne mentionnait quasiment pas les insectes.
* Le néocortex est une structure cérébrale spécifiques des mammifères, constituant la partie externe du cerveau.
Une publication de 2016 par un neurobiologiste et un philosophe spécialiste des neurosciences présentait une revue de la littérature scientifique relative aux analogies fonctionnelles entre le cerveau des insectes et celui des animaux vertébrés (Klein & Barron, 2016). Sur cette base, les auteurs concluaient en faveur de l’idée que les insectes seraient dotés d’une conscience phénoménale leur donnant accès à des états mentaux subjectifs. Cela tiendrait notamment aux caractéristiques d’un ensemble de structures cérébrales des insectes appelé « corps pédonculés » (mushroom bodies en anglais).
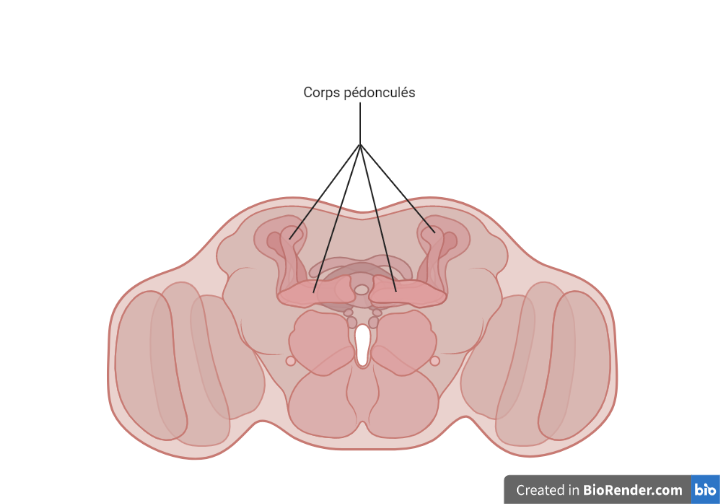
Le rapport de 2017 de l’Inrae sur la conscience faisait part d’une certaine incertitude sur le cas des insectes, tout en laissant la porte ouverte. Les auteurs indiquaient ainsi que « les connaissances actuelles n’excluent cependant pas la possibilité d’une expérience subjective chez les insectes ». En évoquant des travaux sur la neurobiologie et le comportement des abeilles, ils commentaient de la manière suivante : « Ces processus sont totalement cohérents avec les concepts actuels de conscience chez les humains et les autres animaux. »
La conscience de soi renvoie à un niveau de conscience allant au-delà de la simple conscience phénoménale. Certains animaux peuvent être dotés de conscience phénoménale sans forcément être dotés de conscience d’eux-mêmes. Le test du miroir repose sur un protocole permettant d’objectiver si un animal reconnaît sa propre réflexion dans un miroir comme étant lui-même. Il compte parmi la diversité d’outils utilisés par les éthologues pour évaluer la conscience de soi chez les animaux. L’interprétation des résultats de ce test implique un certain nombre de subtilités (notamment l’échec au test du miroir n’est pas considéré comme une preuve de l’absence de conscience de soi).
Lire aussi : Le poisson, le miroir et la conscience de soi (n° 99)
Des travaux préliminaires ont tenté d’étudier la reconnaissance de soi chez les guêpes. Leurs résultats n’ont pas permis d’aboutir à des conclusions claires. En revanche, en 2015, une étude a fait passer le test du miroir à trois espèces de fourmi (Cammaerts Tricot & Cammaerts, 2015). Les auteurs ont comparé leur comportement dans une multitude de situations : fourmis isolées sans miroir, fourmis isolées face à un miroir, fourmis en groupe face à un miroir, fourmis isolées dans une loge où un congénère est visible à travers une vitre, fourmis non marquées, fourmis marquées avec un point bleu visible lorsqu’elles s’observent dans un miroir, fourmis marquées avec un point bleu derrière la tête invisible même lorsqu’elles s’observent dans un miroir, fourmis marquées avec un point de la même couleur que le reste de leur corps, fourmis non marquées exposées à des congénères marqués, fourmis marquées exposées à des congénères marqués, etc.
Les résultats pointent très clairement dans le sens d’une capacité des fourmis à se reconnaître dans un miroir. Cependant, les fourmis juvéniles semblent, elles, ne pas posséder cette capacité. Les auteurs restent malgré tout prudents sur l’interprétation en termes de conscience de soi du fait qu’il s’agit de fourmis[1]. Mais de tels résultats sont habituellement interprétés comme des signes de conscience de soi. L’étude de 2015 a cependant fait l’objet de critiques considérant que la méthodologie ne permet pas d’éliminer certaines hypothèses alternatives (Gallup & Anderson, 2018).
III – Les insectes ressentent-ils la douleur ?
a) Comment déterminer l’existence de douleur chez les animaux ?
Selon l’Association internationale pour l’étude de la douleur (IASP), la douleur se définit comme « une expérience sensorielle et émotionnelle désagréable liée à, ou ressemblant à l’expérience associée à une lésion tissulaire existante ou potentielle ». Elle se distingue de la nociception, définie comme le « processus neural d’encodage d’un stimulus nocif ». Dans l’état actuel des connaissances, la nociception semble être un prérequis à la douleur. Des processus nociceptifs peuvent avoir lieu sans aboutir à l’émergence d’une sensation de douleur. L’activation des nocicepteurs (cellules nerveuses qui détectent les stimuli nocifs), puis des neurones relayant le message le long de la moelle épinière jusqu’au cerveau d’une personne inconsciente blessée est un bon exemple de nociception sans douleur associée.
Lire aussi : Douleur des poissons : va-t-on continuer à noyer le poisson ? (n° 104)
Différents cadres théorico-méthodologiques, en partie influencés par des travaux issus de la philosophie de l’esprit et l’épistémologie, ont été proposés pour procéder à l’évaluation de la capacité à ressentir la douleur. Les auteurs du rapport de la London School of Economics sur les crustacés et les céphalopodes ont proposé un modèle basé sur les huit critères suivants, repris par les auteurs de la récente revue de littérature sur les insectes :
- Nocicepteurs : l’animal possède des récepteurs sensibles aux stimuli nocifs.
- Régions cérébrales intégratives : l’animal possède des structures cérébrales capables d’intégrer les informations issues de différentes sources sensorielles.
- Nociception intégrée : l’animal possède des voies neuronales qui connectent les nocicepteurs aux régions cérébrales intégratives.
- Analgésie : la réponse comportementale de l’animal face à des stimuli nocifs est modulée par des composants chimiques affectant le système nerveux, qu’ils soient endogènes (naturellement produits par l’organisme) ou exogènes (non produits par l’organisme).
- Compromis motivationnels : l’animal réalise des compromis motivationnels dans lesquels la valeur négative des stimuli nocifs ou menaçants est mise en balance face à la valeur positive de l’opportunité d’accéder à une récompense, ce qui donne lieu à une flexibilité dans la prise de décision. Suffisamment de flexibilité doit être démontrée pour indiquer un traitement centralisé et intégratif de l’information impliquant une évaluation bénéfice-risque. La validation de ce critère démontre que la réponse comportementale de l’animal aux stimuli nocifs n’est pas un simple réflexe.
- Auto-protection flexible : l’animal montre des comportements d’auto-protection flexibles (ex : apaiser les blessures en se frottant, se léchant, s’auto-toilettant ou en cachant/évitant d’utiliser la zone endolorie (ex : boiterie) d’une nature qui implique probablement une représentation mentale de la localisation corporelle des stimuli nocifs (voir schémas 2 et 3).
- Apprentissage associatif : l’animal apprend par renforcement à associer les stimuli nocifs à des stimuli neutres (conditionnement Pavlovien), et/ou des nouvelles manières d’éviter les stimuli nocifs.
- Préférence pour l’analgésie : l’animal montre qu’il valorise l’accès à un analgésique ou un anesthésique lorsqu’il est blessé. Cela peut se manifester de différentes manières : auto-administration d’analgésique en cas de blessure, préférence pour les endroits où l’animal a pu accéder à des analgésiques par le passé lorsqu’il est blessé, priorisation de l’accès à l’analgésie par rapport à d’autres ressources (nourriture, abris) en cas de blessure.
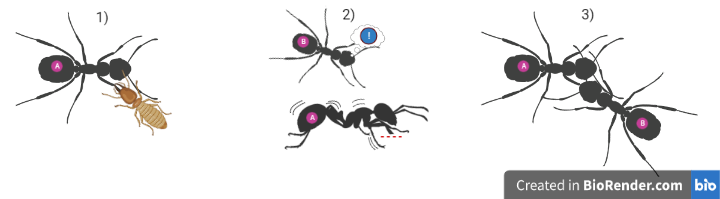
L’originalité de ce modèle tient au fait qu’en plus de proposer une liste de critères, il propose un cadre standardisé quant à la manière de les interpréter. Ainsi, pour chaque critère, pour un groupe d’animaux donné, on attribue un niveau de certitude allant de 1) « certitude très élevée » à 5) « certitude très basse » pour juger de manière graduée si ce critère est validé ou non. On distingue dans cette démarche l’« absence de preuve » (ex : aucune étude n’a été réalisée), de la « preuve de l’absence »[2] (ex : des études ont démontré que le critère est invalidé). Puis on agrège ces informations pour classer les animaux étudiés dans différentes catégories en fonction du niveau de preuve relatif à leur potentielle sensibilité à la douleur. Les auteurs distinguent les catégories suivantes :
- Très haut niveau de preuve : validation de 7 à 8 critères avec un niveau de certitude très élevé ou élevé.
- Fort niveau de preuve : validation de 5 à 6 critères avec un niveau de certitude très élevé ou élevé.
- Niveau substantiel de preuve : validation de 3 à 4 critères avec un niveau de certitude très élevé ou élevé.
- Quelques éléments de preuve : validation de 2 à 3 critères avec un niveau de certitude très élevé ou élevé.
- Sensibilité à la douleur inconnue ou improbable : validation de 0 à 1 critère avec un niveau de certitude très élevé ou élevé. Si la validation des autres critères est inconnue (absence de preuve) et non invalidée (preuve de l’absence), la sensibilité à la douleur est jugée « inconnue ». Si des travaux scientifiques de haute qualité montrent que les autres critères sont invalidés (preuve de l’absence) et non simplement inconnus (absence de preuve), la sensibilité à la douleur est jugée « improbable ».
Les auteurs de la nouvelle revue de littérature sur les insectes émettent une critique de ce modèle bien qu’ils y aient recours : les 8 critères utilisés sont considérés sur un même plan en terme d’interprétation, alors même que certains critères, notamment les compromis motivationnels, pourraient être vus comme des preuves plus fortes que d’autres critères moins évocateurs, tels que la simple présence de nocicepteurs.
b) La douleur chez les insectes
La nouvelle revue de littérature de 2022 s’est concentrée sur 6 grands ordres parmi les insectes : les blattodés (cafards, termites), les coléoptères (scarabées), les diptères (mouches, moustiques), les hyménoptères (abeilles, guêpes, fourmis), les lépidoptères (papillons, mites, vers à soie), et les orthoptères (criquets, sauterelles), en distinguant les adultes des juvéniles (dont les stades larvaires pour les espèces avec métamorphose. Dès lors qu’un critère est validé chez au moins une espèce d’un ordre, cela compte comme un critère validé pour l’ordre entier. Il s’agit d’un raisonnement généralisant par analogie, étant donné qu’il est impossible d’étudier toutes les espèces de chacun de ces ordres. Sur la base de plus de 350 études, les auteurs sont parvenus aux conclusions suivantes :
| Ordre taxonomique | Niveau de preuve de la sensibilité à la douleur pour les adultes | Niveau de preuve de la sensibilité à la douleur pour les juvéniles |
| Blattodés | 6 critères satisfaits : fort niveau de preuve | 3 critères satisfaits : niveau de preuve substantiel |
| Coléoptères | 2 critères satisfaits : quelques éléments de preuve | 1 critère satisfait : sensibilité inconnue |
| Diptères | 6 critères satisfaits : fort niveau de preuve | Juvéniles tardifs : 4 critères satisfaits: niveau de preuve substantiel Juvéniles précoces : 3 critères satisfaits: niveau de preuve substantiel |
| Hyménoptères | 4 critères satisfaits : niveau de preuve substantiel | 1 critère satisfait : sensibilité inconnue |
| Lépidoptères | 3 critères satisfaits : niveau de preuve substantiel | Juvéniles tardifs : 3 critères satisfaits : niveau de preuve substantiel Juvéniles précoces : 1 critère satisfait : sensibilité inconnue |
| Orthoptères | 3 critères satisfaits : niveau de preuve substantiel | 2 critères satisfaits : quelques éléments de preuve |
Les auteurs précisent par ailleurs qu’ils n’ont trouvé « aucune preuve robuste qu’aucun de ces ordres n’échoue à satisfaire l’un des critères ». Ils entendent par là que lorsque des critères sont considérés comme insatisfaits, cela tient à une absence de preuve (absence d’études), et non à une preuve de l’absence. Cela sous-entend que pour les ordres ou les stades de vie pour lesquels peu de critères sont satisfaits à ce jour, il faut plutôt considérer que leur sensibilité à la douleur est inconnue plutôt qu’improbable. Ils soulignent notamment la pauvreté des connaissances actuelles concernant les stades juvéniles.
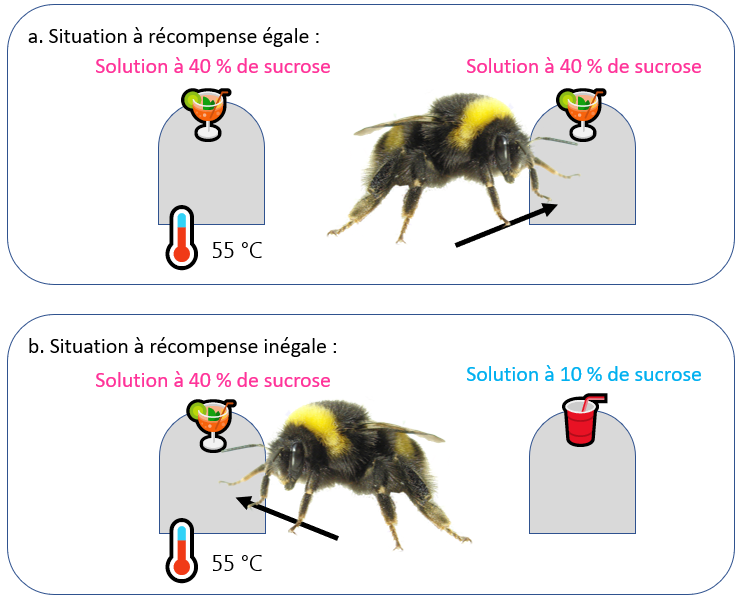
Une expérience démontrant un compromis motivationnel flexible chez les bourdons est particulièrement parlante. Des bourdons ont été entraînés à utiliser plusieurs distributeurs d’eau sucrée de couleur différente (Gibbons et al., 2022). Certains de ces distributeurs étaient chauffés à des températures présumées légèrement douloureuses. Au début de l’expérience, les différents distributeurs proposaient tous de l’eau avec une concentration de sucre de 40 %. Les bourdons évitaient alors les distributeurs chauffés, et préféraient les distributeurs non chauffés. Plus tard, la concentration en sucre des distributeurs non chauffés a été fortement diminuée. Les seuls distributeurs proposant encore une concentration de sucre élevée étaient alors les distributeurs chauffés. Dans ce contexte, les bourdons se mettaient à utiliser les distributeurs chauffés. Cela montre que lorsqu’ils peuvent accéder à de l’eau très sucrée sans se brûler, les bourdons ne voient pas d’intérêt à prendre le risque de se brûler sur les distributeurs chauffés. Cependant, lorsque cette option leur est retirée, ils sont alors capables de se dire que le bénéfice de l’accès à l’eau très sucrée peut valoir la peine d’endurer une légère brûlure. Cela montre une capacité à évaluer une balance bénéfice-risque de manière flexible. Cela indique que leur évitement initial des distributeurs chauffés est une décision réfléchie, signalant un traitement intégré de l’information, et non un simple réflexe.
Les conclusions de cette dernière revue sont beaucoup plus affirmatives de l’idée que les insectes ressentiraient la douleur comparée à la plupart des publications précédentes à ce sujet. Celles-ci, souvent basées sur une approche centrée sur la recherche de structures cérébrales similaires à celles des mammifères ou plus largement des vertébrés, concluaient que les insectes ne ressentent pas la douleur.
En 2005, dans le cadre d’un rapport destiné à aider les décideurs à encadrer l’expérimentation animale, l’Autorité européenne de sécurité des aliments (EFSA) a produit un rapport sur la sensibilité à la douleur chez les invertébrés. [Notons par ailleurs que, dès 2005, l’EFSA considérait déjà que les crustacés et céphalopodes ressentent probablement la douleur.] L’EFSA classifiait alors divers taxons dans trois catégories : les animaux qui ressentent probablement la douleur, les animaux qui ne ressentent probablement pas la douleur, et ceux pour lesquels on ne pouvait pas encore se prononcer. Dans ce rapport, l’EFSA avait classé les insectes sociaux dans la catégorie « incertaine », mais avait classé les autres insectes dans la catégorie des animaux qui ne ressentent probablement pas la douleur.
En 2014 fut publiée une revue de littérature sur la sensibilité à la douleur chez une diversité d’animaux basée sur l’analyse de 17 critères (Sneddon et al., 2014). Elle considérait que, concernant les insectes, 8 critères étaient validés, 7 demeuraient irrésolus (absence de preuve), et 2 étaient invalidés (preuves de l’absence). Le premier critère considéré comme invalidé renvoyait à des travaux montrant une absence de boiterie (critères de comportement d’auto-protection) chez certains insectes pourtant blessés aux pattes. Le second renvoyait et à des travaux montrant que des insectes sévèrement blessés peuvent parfois continuer à s’adonner à d’autres activités sans prioriser la gestion de la douleur d’autre part (critère de réponse prioritaire accordée aux stimuli nocifs par rapport à d’autres stimuli). C’est en particulier le cas dans le contexte des activités sexuelles, avec les mâles des mantes religieuses qui continuent l’accouplement alors qu’ils sont en train d’être dévorés vivants par la femelle. Les auteurs de la revue de 2014 faisaient alors part d’une certaine incertitude dans leurs conclusions sur la sensibilité à la douleur des insectes.
Le critère de réponse prioritaire accordée aux stimuli nocifs par rapport à d’autres stimuli ne figure pas parmi les critères du modèle d’analyse de la revue de 2022. Quant à l’absence de boiterie des insectes blessés aux pattes, les auteurs de la revue de 2022 ont attribué un niveau de certitude très bas à cette observation. Cela est lié à la méthodologie de l’étude, qui n’a pas procédé à des mesures quantifiées. Ces éléments expliquent la divergence entre la publication de 2014 et celle de 2022 quant à l’invalidation de certains critères.
IV – Les insectes ressentent-ils le plaisir ?
Contrairement à la douleur, il ne semble pas exister d’étude centrée exclusivement sur l’évaluation de la capacité des insectes à ressentir le plaisir. Ce concept semble d’ailleurs moins précisément défini que la douleur.
Chez les animaux vertébrés dont l’humain, on appelle « système de récompense » un ensemble de structures cérébrales (notamment le noyau accumbens, l’aire tegmentale ventrale et le pallidum ventral) qui s’activent généralement lors d’expériences agréables telles que la consommation alimentaire, les rapports sexuels et la prise de drogues (Berridge & Kringelbach, 2008). L’activation de ces structures repose généralement sur certains types de neurotransmetteurs/neurohormones précis comme la dopamine, les opioïdes et les endorphines. Le système de récompense implique à la fois des structures corticales* et sous-corticales. Les zones sous-corticales du cerveau sont généralement vues comme plus anciennes sur le plan évolutif et davantage similaires entre les différents taxons d’animaux. Le système de récompense est impliqué dans le plaisir, mais aussi plus largement dans les processus liés à la motivation tels que l’envie / le désir (craving). Sur le plan comportemental, on considère que le fait qu’un stimulus puisse être utilisé comme une récompense dans le cadre d’un apprentissage, et que l’animal fasse preuve d’une motivation à y accéder sont des indicateurs de sa valeur hédonique positive – c’est-à-dire sa valeur en terme de plaisir ressenti.
* Une zone « corticale » est une zone du cortex, partie externe du cerveau. Une zone « sous-corticale » est une zone située plus profondément à l’intérieur du cerveau, en dessous du cortex.
Par rapport à la douleur, les récepteurs sensoriels impliqués dans le plaisir semblent moins spécifiques au plaisir. Le plaisir physique semble ainsi résulter du mariage de l’activation d’une diversité de récepteurs sensoriels non spécifiques et de l’activation du système de récompense. Par rapport à la douleur, qui est fondamentalement une expérience sensorielle, le plaisir pourrait parfois être déconnecté d’une sensation physique. En effet, le système de récompense peut s’activer en réponse aux interactions sociales positives chez les espèces sociales.
Il est intéressant de noter qu’aucun néologisme équivalent à « nociception » dans le contexte du plaisir n’est réellement en usage répandu. On aurait pu pourtant imaginer une distinction entre « hédoception » – processus neural d’encodage d’un stimulus hédonique – et « plaisir » au sens de ressenti subjectif sensoriel et émotionnel agréable. Bien qu’ils n’utilisent pas ces termes, certains neurobiologistes du plaisir considèrent bien qu’une distinction de ce type existe. Ils soulignent qu’il est possible d’observer une activation du système de récompense en réponse à un stimulus hédonique sans nécessairement que cela soit associé à un ressenti subjectif conscient de plaisir. Ce serait notamment le cas lors de l’exposition à des stimuli potentiellement agréables subliminaux (allant trop vite pour que l’on en ait conscience). La jouissance consciente nécessiterait que l’activation du système de récompense soit combinée à d’autres processus neuraux d’intégration* additionnels.
*Par processus intégratifs on entend les processus permettant d’associer ensemble les informations issues de plusieurs sources sensorielles différentes (après un premier traitement des informations issues de chacun des sens de manière séparée) au sein du cerveau et de les traiter ensemble sur le plan cognitif.
Si aucune étude ne tente de savoir si les insectes ressentent le plaisir, un ensemble de données issues de travaux portant sur d’autres sujets peut tout de même être utile dans ce contexte. Notamment, le système nerveux des insectes semble bien doté d’un système de récompense. Il présente certaines homologies structurelles et fonctionnelles avec celui des animaux vertébrés (Søvik et al., 2015 ; Perry & Barron, 2013). Cet aspect est particulièrement étudié chez les drosophiles et les abeilles. On constate toutefois aussi des différences, notamment chez certains insectes qui sont dépourvus de récepteurs aux opioïdes.
D’autre part, sur le plan comportemental, les insectes sont tout à fait capables de réaliser des apprentissages basés sur des récompenses alimentaires telles que l’eau sucrée. De plus, les insectes sont parfois utilisés comme modèles d’expérimentation animale pour étudier les addictions à diverses drogues, notamment l’alcool (Dvořáček & Kodrík, 2022 ; Søvik & Barron, 2013). Ces éléments peuvent plaider en faveur de l’idée qu’il n’est pas déraisonnable de penser que les insectes pourraient bien ressentir le plaisir. Toutefois, si les travaux sur la douleur chez les insectes tiennent compte de la distinction entre douleur et nociception, une distinction équivalente n’est pas faite dans les travaux sur l’apprentissage par récompense et sur les drogues ou addictions qui les concerne. En effet, le but de ces études n’est pas d’étudier le plaisir chez les insectes. Aussi, il est difficile de dire si l’activation du système de récompense des insectes relève simplement de l’« hédoception » ou est génératrice d’un ressenti subjectif de plaisir.
V – Les insectes ressentent-ils des émotions ?
Il existe plus de 90 définitions différentes du mot « émotion » (Kleinginna & Kleinginna, 1981). On considère généralement qu’il s’agit de schémas généraux (patterns) de réaction psychologique à la perception d’une situation caractérisés par des dimensions physiologiques (ex : variations du rythme cardiaque), cognitives (ex : pensées de type « je suis content », biais cognitifs spécifiques), subjectives (ressenti conscient), comportementales (ex : expressions faciales) et par une valence positive ou négative, ainsi qu’une valeur adaptative sur le plan évolutif.
L’aspect cognitif se caractérise notamment par le fait que les émotions positives ont tendance à induire un biais optimiste tandis que les émotions négatives ont tendance à induire un biais pessimiste (Baciadonna & McElligott, 2023 ; Mendl et al., 2009). La présence de ces biais cognitifs émotionnels optimistes et pessimistes est bien établie chez divers animaux non-humains. On appelle « test du biais de jugement » le protocole utilisé par les éthologues pour évaluer ces biais pessimistes ou optimistes en réponse à des stimuli à valence émotionnelle. Ce test est couramment utilisé comme un indicateur de l’état émotionnel des animaux, notamment dans le cadre des recherches sur le bien-être des animaux d’élevage.
Lire aussi : Les poissons ont-ils des chagrins d’amour ? (n° 104)
Quelques études ont démontré l’existence de biais optimistes ou pessimistes lors de tests de biais de jugement chez les insectes. Ainsi, les bourdons exposés à une récompense alimentaire sucrée de manière inattendue présentent un biais de jugement optimiste (Solvi et al., 2016 ; Plowright, 2017). Celui-ci disparaît si on leur donne au préalable une drogue bloquant l’activité de la dopamine, neurotransmetteur impliqué dans le plaisir et les émotions positives. À l’inverse, les abeilles et les drosophiles préalablement secouées dans un petit conteneur montrent un biais de jugement pessimiste (Schlüns et al., 2017 ; Deakin et al., 2018).
L’interprétation de ces quelques études est cependant sujette à débat (Barrachi et al., 2017). D’autre part, une étude récente a montré que lorsqu’on leur donne accès à de petites balles, les bourdons interagissent spontanément avec elles, d’une manière qui correspond aux critères de certaines définitions du jeu chez les animaux (Dona et al., 2022). Notamment, l’accès à ces balles-jouet peut être utilisé avec succès comme une récompense dans le cadre d’un apprentissage. Cela suggère que le jeu avec une balle pourrait être une source de ressenti positif, peut-être émotionnel, chez les bourdons.
Ces études peuvent plaider en faveur de l’idée qu’au moins certains insectes pourraient bien être capables de ressentir des émotions positives ou négatives. Dans l’ensemble, si les rares études disponibles penchent en faveur de capacités émotionnelles chez les insectes, leurs éventuelles émotions ne sont pas encore aussi bien établies que pour les animaux vertébrés. Il conviendrait idéalement d’évaluer si les dimensions autres que cognitives sont aussi à l’œuvre (et de manière simultanée) dans les situations où l’on soupçonne que les insectes ressentent des émotions.
VI – Conclusion
Dans l’ensemble, les études disponibles ont tendance à converger en faveur de l’idée qu’au moins certains insectes sont probablement bien sentients. La sensibilité à la douleur est l’aspect de leur sentience le plus étudié, les études sur leurs niveaux de conscience, leur sensibilité au plaisir et leurs capacités émotionnelles étant plus préliminaires. La sentience des insectes adultes semble davantage établie que celles des stades juvéniles. Du fait de l’extrême diversité du groupe des insectes, il n’est pas impossible que certaines espèces puissent être sentientes et que d’autres ne le soient pas. Et beaucoup de zones d’ombres subsistent.
Ces travaux sur la sentience des insectes ont d’énormes implications éthiques. Ils alimenteront nécessairement les débats relatifs à leur éventuelle protection juridique, à l’effondrement de leurs populations sauvages, et à leur utilisation à des fins scientifiques. En effet, les céphalopodes sont actuellement les seuls invertébrés protégés par le droit européen de l’expérimentation animale. Les chercheurs indépendants de l’ONG Wild Animal Initiative réfléchissent déjà à la question de la réduction des douleurs infligées aux insectes dans le cadre de l’usage des insecticides. Ils proposent de privilégier les molécules les moins génératrices de souffrances lorsque c’est possible.
Le rapide développement actuel de l’élevage d’insectes – essentiellement pour les utiliser pour alimenter les animaux d’élevage – est sûrement l’enjeu le plus épineux. Il fait dire à Lars Chittka, co-auteur de la récente revue de littérature sur la douleur : « Un moyen simple et évident de minimiser les risques éthiques en matière de bien-être animal serait de ne pas élever d’animaux du tout. Mais l’élevage d’insectes connaît actuellement une croissance exponentielle. Nous devons nous assurer que des mesures soient prises pour éviter que les mêmes erreurs que celles commises dans l’élevage en batterie des volailles, par exemple, ne se répètent encore et encore. »
Lire aussi: « Quelle qualité de vie pour les insectes d’élevage? », revue n°120
Preuve de la montée des préoccupations pour les invertébrés, le lancement d’une organisation spécifiquement dédiée à l’amélioration des conditions d’élevage des insectes, l’Insect Institute, a été annoncée récemment aux États-Unis.
Gautier Riberolles
Pour aller plus loin sur le concept de sentience et son évaluation :
- Projet Méduse rassemblant des ressources scientifiques sur la sentience et des ressources philosophiques sur le sentientisme.
- Conférence de Victor Duran Le Peuch aux Estivales de la Question Animale 2022 : « Comment déterminer scientifiquement qui est sentient ? ».
1. Entre autres observations, cette étude fait ressortir que des fourmis sur qui l’on appose une marque de couleur bleue au niveau de la tête se mettent à la nettoyer si elles sont face à un miroir, mais ne le font pas si elles ne sont pas face à un miroir. L’absence de tentative de nettoyer la marque chez les fourmis qui ne sont pas face à un miroir constitue un groupe contrôle indispensable. Or, on reproche aux auteurs de l’étude de ne pas avoir indiqué le temps écoulé entre le marquage et le moment où le comportement des fourmis était observé. Si les fourmis du groupe contrôle sans miroir touchaient aussi leur marque pendant cette durée avant le début des observations, on pourrait considérer que les tentatives des fourmis en présence d’un miroir de retirer la marque ne sont pas une preuve suffisante de reconnaissance de soi. On reproche également un manque de précision sur la méthodologie d’observation du comportement utilisée et le fait qu’aucun test d’accord inter-observateur (indice permettant de vérifier la fiabilité des observations faites par différents observateurs) n’ait été réalisé. L’étude de 2015 montre que lorsqu’on remplace la marque bleue par une marque d’une couleur similaire à celle du corps des fourmis, les signes comportementaux de reconnaissance de soi lors de l’exposition à un miroir disparaissent, potentiellement car la marque n’est alors plus visible dans la réflexion du miroir. Les critiques mettent en avant l’hypothèse selon laquelle, plutôt qu’un effet lié aux couleurs, cette différence de comportement pourrait aussi s’expliquer par une différence d’odeur entre les peintures utilisées. Cela n’expliquerait pas, toutefois, la différence de comportement des fourmis marquées à la peinture bleue selon qu’elles soient face à un miroir ou en absence de miroir, ni le fait que lorsque la même peinture bleue est utilisée pour appliquer une marque à l’arrière de la tête (localisation invisible dans la réflexion) les fourmis ne tentent pas d’y toucher, qu’elles soient ou non face à un miroir, alors qu’elles tentent d’enlever la marque bleue lorsqu’elle est disposée de manière visible dans leur réflexion. Enfin, les critiques rappellent que l’on ignore, avant-même de parler de reconnaissance de soi, si les fourmis ont ou non la capacité à discriminer et reconnaître individuellement leurs congénères exclusivement par la vision. En effet, les fourmis ont plutôt une mauvaise vision, leur sens principal étant plutôt la perception des stimuli chimiques. Cela rend leur éventuel succès au test du miroir très étonnant. En effet, des travaux montrent que les animaux pour qui la vision n’est pas une modalité sensorielle privilégiée ont tendance à échouer ce test, mais peuvent parfois réussir des tests de conscience de soi basés sur d’autres modalités sensorielles (ex : odorat). Retour
2. La « preuve de l’absence » est beaucoup plus compliquée à apporter que la « preuve de la présence » en science. En pratique, cela renvoie surtout au cas où une étude essaierait de tester si un critère de sensibilité à la douleur est validé mais échouerait à y parvenir. Certains philosophes des sciences considèrent qu’une véritable preuve d’absence est conceptuellement impossible. Ils considèrent alors qu’il faut plutôt réfléchir selon la maxime « pas de preuve = pas de raison de croire ». Cette maxime n’est pas totalement consensuelle pour autant, dans la mesure ou d’autres approches en philosophie des sciences (bayésianisme) mettent en avant la valeur scientifique des hypothèses par défaut selon un raisonnement probabiliste basé sur les connaissances précédentes, même en cas d’incertitude. Dans le cas de la douleur, de telles hypothèses par défaut peuvent notamment mettre en avant la proximité évolutive des insectes avec les crustacés décapodes – dont la sensibilité à la douleur est davantage établie – et des raisonnements théoriques sur les mécanismes évolutifs derrière l’apparition de la sensibilité à la douleur. Retour

