Le 30 septembre dernier, la brillante biologiste Victoria Anne Braithwaite est décédée. Elle fut pionnière dans l’étude de la douleur chez les poissons en cosignant deux articles (1,2) en 2003 avec Lynne Sneddon et Michael Gentle. Ces publications furent les premières d’une série ouvrant le débat scientifique et éthique sur la douleur des poissons. En hommage à une dame de science, nous vous proposons un aperçu des connaissances scientifiques et des questions morales que suscitent la douleur chez ces porteurs de branchies et de nageoires.

Qu’est-ce que la douleur ?
L’association internationale pour l’étude de la douleur (IASP) propose la définition suivante : « une expérience sensorielle et émotionnelle désagréable, associée à des dommages tissulaires réels ou potentiels, ou rapportés comme tels ». Cette définition s’accompagne d’une longue note qui précise que « l’incapacité à communiquer verbalement ne réfute pas la possibilité qu’un individu fasse l’expérience de la douleur et requiert un traitement pour soulager sa douleur ». Cette précision est d’une importance toute particulière en ce qui concerne les nourrissons, les personnes en situation de handicap et les animaux.
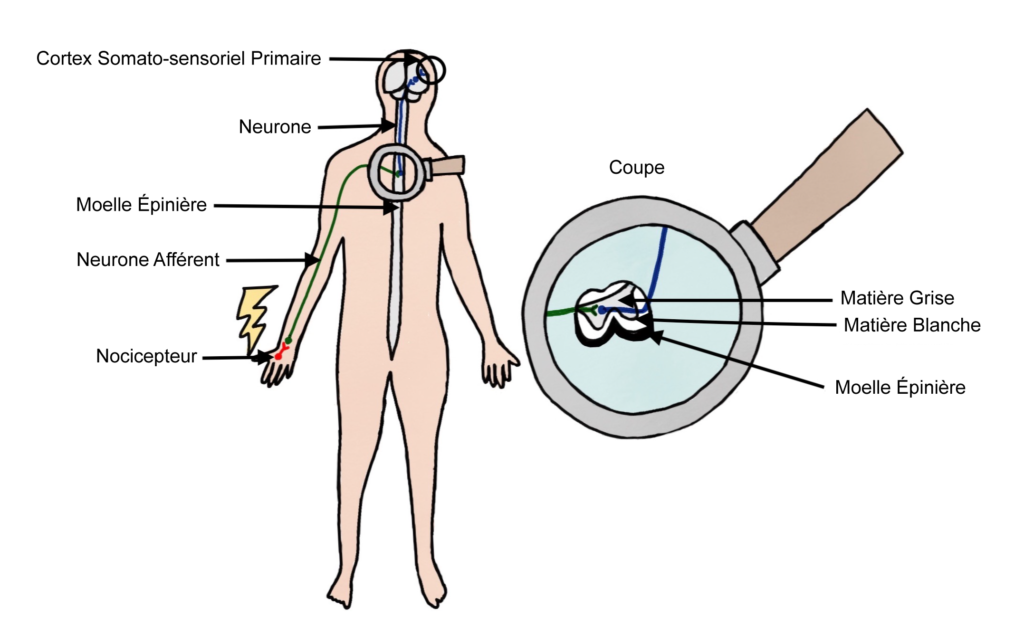
La douleur est avant tout une sensation subjective. Elle est à distinguer de la nociception qui renvoie au « processus neural d’encodage d’un stimulus nocif », lequel n’implique pas nécessairement l’expérience mentale de la douleur. Le système repose sur les nocicepteurs : des cellules nerveuses spécialisées dans la détection des stimuli nocifs et situées dans le corps (peau, muscles, articulations…). Ces cellules s’activent en envoyant un message électrique lorsqu’elles détectent un signal physique dangereux comme une température trop élevée, une pression mécanique trop forte, un déchirement des tissus ou une substance chimique agressive. Par « processus neural d’encodage d’un stimulus nocif », on veut dire que les nocicepteurs s’activent en envoyant un signal électrique qui va se propager le long d’une chaîne de cellules nerveuses et éventuellement (mais pas nécessairement) atteindre le cerveau où un réseau de neurones analysera le message reçu. Ce processus de nociception n’entraîne pas obligatoirement de sensation subjective douloureuse. Ainsi, la nociception peut avoir lieu chez des patients humains alors qu’ils sont inconscients, et il a été démontré que l’activation des nocicepteurs n’entraîne pas de douleur tant que le message nerveux n’a pas remonté la moelle épinière puis a été traité par le cerveau. La nociception peut aussi avoir lieu chez des animaux dépourvus de cerveau comme les anémones, sans nécessairement qu’une sensation de douleur soit associée. Chez ces animaux, la nociception s’accompagne souvent de mouvements réflexes protecteurs comme la rétractation des tentacules chez l’anémone. On parle de réflexe, lorsque le signal électrique envoyé par les nocicepteurs se propage non pas vers le cerveau, mais directement vers des cellules nerveuses connectées aux muscles, qui leur donnent l’ordre de réagir en leur envoyant à leur tour un signal électrique.
Comment savoir si un animal ressent la douleur ?
Comment faire la distinction entre les animaux capables de nociception inconsciente sans douleur associée et ceux capables d’éprouver subjectivement la douleur ? La réponse à cette question est à la fois scientifiquement difficile à apporter, et cruciale d’un point de vue éthique.
La plupart des gens reconnaissent intuitivement, sans même se préoccuper de l’existence de preuves, que les humains, les mammifères et les oiseaux peuvent ressentir la douleur. Mais lorsque l’on s’aventure chez des espèces plus éloignées de nous dans l’arbre phylogénétique (l’arbre représentant les relations de parenté entre les différentes branches du vivant au cours de l’évolution), comme les insectes, les méduses, les coquillages, les pieuvres et les poissons : les avis divergent.
L’argument commun est que l’on saurait avec une certitude absolue que les humains souffrent car ils peuvent l’exprimer avec des mots, tandis que le doute est permis pour les autres animaux – ou a minima pour certaines espèces – car ils ne parlent pas. Cette absence de parole empêcherait de conclure sur la présence de douleur chez les animaux, alors que la douleur humaine serait indubitable.
Cependant, le langage ne nous permet pas de faire l’expérience directe à la première personne de ce que ressentent les autres. La douleur fait partie de ce que la philosophie de l’esprit nomme qualia, c’est-à-dire un ressenti subjectif conscient qu’il est impossible, à moins de disposer de dons de télépathie, de partager directement avec autrui. Le mieux que l’on puisse faire est imaginer la sensation que les autres ressentent lorsqu’ils ont mal, à partir de notre expérience personnelle.
Qu’il s’agisse des autres humains ou des animaux, il est impossible d’apporter une preuve définitive et absolue qu’un individu autre que soi-même ressente la douleur. Cependant, nous pouvons utiliser les observations que nous avons faites sur un individu et nous demander si, au vu de ces observations, il semble probable ou improbable que cet individu ressente la douleur. C’est déjà ce que nous faisons en acceptant que les autres humains ressentent la douleur car ils affirment qu’ils ressentent la douleur. Face à l’impossibilité du report verbal de la douleur pour les animaux, nous pouvons utiliser d’autres observations.
La définition la plus commune de la douleur en ce qui concerne les animaux non humains est celle proposée par Zimmerman en 1986 : « une expérience sensorielle désagréable causée par une blessure réelle ou potentielle qui entraîne des réactions végétatives [(c’est-à-dire des modifications physiologiques)] et de protection, résulte en un apprentissage, et peut modifier les comportements spécifiques à l’espèce » .
Lynne Sneddon, qui a obtenu sa thèse sous la direction de Victoria Braithwaite, propose quant à elle deux principes généraux pour identifier la douleur chez les animaux (2, 3). Le premier principe est que l’organisme entier de l’animal (et non seulement les nocicepteurs) doit présenter des réactions aux stimuli potentiellement douloureux qui soient différentes des réactions à des stimuli non douloureux. Par exemple, on va comparer les réactions d’un animal lorsqu’on le touche sans le blesser et lorsqu’on le blesse. Le second principe est que l’animal doit présenter des changements dans les comportements dits motivés (c’est-à-dire les comportements modulés par la recherche d’émotions positives et l’évitement des émotions négatives) après avoir subi un stimulus potentiellement douloureux.
À partir de ces deux principes, combinés à la recherche de structures biologiques qui ressemblent aux structures impliquées dans la douleur humaine, il est possible d’établir une liste de critères à vérifier pour évaluer la capacité d’un animal à ressentir la douleur. Voici un aperçu non exhaustif des critères utilisés par la recherche :
- Présence de nocicepteurs connectés au cerveau.
- Traitement des signaux issus des nocicepteurs par le cerveau, en particulier par des zones liées aux émotions négatives et à la mémoire.
- Les réactions à une stimulation nocive vont au-delà du simple réflexe de retrait.
- Les stimuli nocifs provoquent des réactions physiologiques associées au stress (respiration et rythme cardiaque accélérés, sécrétion d’hormones du stress comme le cortisol).
- Présence de comportements de protection de la zone du corps douloureuse (l’animal évite d’exposer la zone, la lèche, la frotte, la nettoie, il boite pour ne pas utiliser la zone endolorie).
- Présence de récepteurs à une substance analgésique (anti-douleur) sécrétée par l’organisme. Les réponses aux stimuli nocifs sont réduites par l’administration de substances analgésiques.
- L’animal répond aux stimuli douloureux de manière prioritaire par rapport aux stimuli non douloureux (ex : l’animal blessé est moins attentif à des stimuli auxquels il prête attention lorsqu’il n’est pas blessé).
- L’animal est prêt à perdre des ressources ou à s’exposer au danger pour éviter ou soulager une douleur.
- L’animal apprend à éviter les zones ou les objets associés à la douleur. Cet apprentissage dit « aversif » se maintient sur le long-terme si le stimulus douloureux est répété.
- L’animal fait des compromis entre la motivation à éviter la douleur et d’autres motivations (ex : évitement d’un distributeur de nourriture où l’animal a reçu un choc électrique jusqu’à ce que la faim devienne trop forte et pousse l’individu à retourner au distributeur malgré le risque de recevoir un choc électrique).
Aucun de ces critères pris isolément ne permet de trancher de manière absolue. Cependant, il est raisonnable de penser que plus une espèce animale remplit les critères de cette liste, plus il est probable qu’elle soit capable de ressentir la douleur. Si une espèce valide tous les critères, l’hypothèse selon laquelle l’animal ne ressentirait pas la douleur devient plus difficile à soutenir que l’hypothèse plus parcimonieuse selon laquelle l’animal ressent la douleur. Si une espèce possède des nocicepteurs et a des réactions comportementales qui nous rappellent les nôtres face à la douleur, le plus simple est de supposer que cette espèce et la nôtre ont hérité cette sensibilité d’un ancêtre commun. Il est aussi envisageable que cette capacité ait été acquise indépendamment, de façon convergente, comme l’apparition d’ailes chez les oiseaux et les chauves-souris par des chemins évolutifs différents. Soutenir que l’espèce ne ressent pas la douleur malgré la validation des critères demande d’expliquer pourquoi et comment l’évolution a abouti à ces caractéristiques anatomiques et comportementales sans qu’elles soient liées à une perception de la douleur. Cela fait appel à de nombreuses inconnues en termes de processus évolutifs.
La douleur chez les poissons
Dans une revue de 2015, Lynne Sneddon avance que tous les critères susmentionnés ont été validés pour le groupe des poissons téléostéens. Ce groupe comprend 96 % des espèces de poisson recensées, dont la quasi-totalité des espèces utilisées pour la consommation. Il exclut cependant les requins et les raies, très éloignées dans l‘arbre phylogénétique (représentation des liens de parentés entre les différentes branches du vivant) . Voici un aperçu des résultats les plus convaincants.
Avant 2002, il avait été affirmé que les poissons ne pouvaient pas ressentir la douleur car on n’avait pas confirmé chez eux la présence de nocicepteurs. Mais en 2002 et en 2003, on identifia pour la première fois des nocicepteurs chez la truite arc-en-ciel, particulièrement nombreux au niveau des lèvres (3). Il fut par la suite démontré chez les carpes, les poissons rouges, les truites et les saumons, que les stimuli potentiellement douloureux engendraient une activité dans des zones du cerveau différentes de l’activité observée en réaction à des stimuli non douloureux (2).
L’injection de venin d’abeille ou de vinaigre dans les lèvres chez les truites et les poissons zèbre provoque des réactions de stress comme la sécrétion de cortisol et l’augmentation du rythme de la respiration (2). Les poissons évitent alors de manger pendant plusieurs heures et vont jusqu’à se frotter les lèvres contre le substrat ou les vitres de l’aquarium (2, 4). Or, ces réactions sont minimes voire inexistantes lorsque les poissons sont traités par injection de liquide physiologique indolore plutôt qu’avec des substances agressives, et elles sont réduites lorsqu’ils reçoivent des analgésiques .
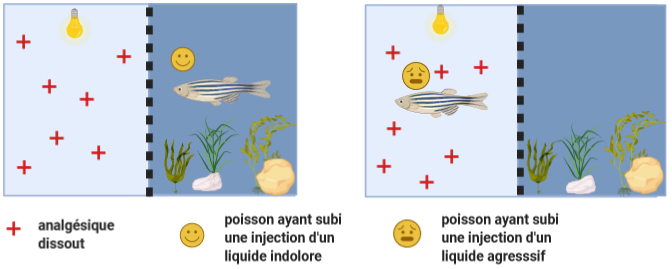
Des compromis entre différentes motivations sont également observés. Ainsi, les truites et les poissons rouges évitent une zone de nourrissage où ils ont reçu des chocs électriques (3). Ces poissons se retiennent alors de manger jusqu’à 3 jours pour éviter d’entrer de nouveau dans la zone où ils risquent de recevoir un choc électrique (3). Ces réponses sont diminuées en cas d’injection de morphine connue pour son action analgésique (3). Plus frappants encore sont les résultats d’une étude de 2013 sur les poissons zèbres. Lorsqu’on leur laisse le choix entre une zone sombre et enrichie par de la végétation et du substrat, et une zone lumineuse et nue, ces animaux choisissent systématiquement la zone enrichie, probablement parce qu’elle leur procure une protection rassurante face à des prédateurs potentiels. Si l’on injecte du vinaigre ou du liquide physiologique indolore à ces poissons, ils continuent de manifester leur préférence pour la zone sombre dans les deux cas. Cependant, si l’on dissout un anti-douleur dans l’eau de la zone lumineuse et nue, les individus qui ont subi une injection de vinaigre accepteront de s’exposer au danger dans la zone lumineuse pour réduire leur douleur en nageant dans l’eau de cette zone. En revanche, les individus à qui l’on a injecté du liquide physiologique indolore maintiennent leur préférence pour la zone sombre et enrichie, plus rassurante. On peut interpréter ces résultats comme la démonstration que les poissons zèbres acceptent de payer un coût (ils s’exposent au danger) pour pouvoir réduire leur douleur.
En temps normal, les truites sont craintives lorsqu’elles rencontrent un nouvel objet dans leur environnement, comme une tour de briques de plastique, et l’évitent au maximum : on parle de néophobie. Or, après une injection douloureuse de vinaigre, cette peur des objets nouveaux disparaît. Cependant, si on injecte de la morphine aux animaux pour calmer leur douleur, les truites se montrent craintives face aux objets nouveaux comme à leur habitude. Pour Lynne Sneddon, la douleur perturbe l’attention des animaux pour qui la peur naturelle des objets inconnus est atténuée car ils ont « la tête ailleurs » (2).
Les études à base de compromis entre différentes motivations ou de perturbation de l’attention sont très importantes. En effet, il ne peut pas s’agir ici de réactions de nociception réflexe inconsciente comme chez les anémones. Les situations que nous avons évoquées ont en commun de démontrer que chez ces animaux, la douleur modifie des décisions issues de processus intellectuels complexes comme l’attention ou l’évaluation du danger. Ces comportements ne peuvent pas être des réflexes car ils nécessitent une coordination d’actions différentes et l’utilisation de différentes informations en compétition les unes avec les autres (ex : éviter un choc en risquant la dénutrition, ou bien se nourrir en risquant un choc, etc.) .
Face à l’accumulation des études, Victoria Braithwaite disait : « Il y autant de preuves que les poissons ressentent la douleur qu’il n’y en a pour les mammifères et les oiseaux, et plus qu’il n’y en a pour les nourrissons et les prématurés humains » .
Controverse scientifique ou consensus ?
Pourtant, quelques scientifiques continuent de soutenir que les poissons ne ressentiraient pas la douleur. James D. Rose et Brian Key sont les principaux défenseurs de ce point de vue. Ils avancent principalement deux arguments.
① Le premier argument est que le néocortex, la partie externe du cerveau, est une structure uniquement présente chez les mammifères. Des éléments suggèrent que le néocortex serait le siège de la conscience chez les humains (voir l’article sur la conscience dans le n° 94 de cette revue). Or cette structure est absente chez les poissons, qui seraient donc des êtres sans conscience. Le pallium, la structure cérébrale plus ou moins équivalente au néocortex chez les poissons, ne posséderait pas les caractéristiques structurelles nécessaires au traitement de la douleur chez les humains (5, 6). D’ailleurs l’ablation du pallium chez certains poissons ne les empêche pas de manifester des mouvements de retrait après un choc électrique (5, 6).
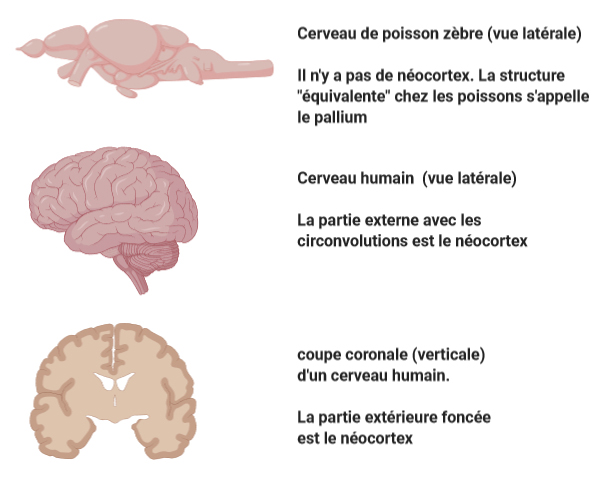
Cet argument de l’absence de néocortex est cependant réfuté par d’autres auteurs qui soutiennent que cette vision est anthropocentrique, car elle exclue la possibilité que des structures cérébrales autres que le néocortex pourraient générer la conscience (7). Cet argument exclue également les oiseaux du cercle des êtres conscients, alors que beaucoup d’études entérinent la présence de niveaux de conscience chez certains oiseaux (7). Enfin, les sciences de l’évolution ont recensé plusieurs fois des preuves que des structures cérébrales apparues récemment dans l’évolution, comme le néocortex, ont progressivement récupéré des fonctions qui étaient par le passé exercées par d’autres structures plus primitives (8). Ainsi, il semble tout à fait possible que les ancêtres des mammifères, à l’époque dépourvus de néocortex, disposaient déjà de structures cérébrales capables de traiter consciemment la douleur, puis que la fonction « traitement de la douleur » ait été prise en charge ensuite progressivement par le néocortex après son apparition, en remplacement des structures primitives (8). La célèbre Déclaration de Cambridge de 2012 sur la conscience – un manifeste de consensus rédigé par un ensemble d’experts en neuroscience – affirme elle aussi que « l’absence d’un néocortex ne semble pas empêcher un organisme de connaître des états affectifs ».
② Le second argument contre l’existence de la douleur chez les poissons tient au fait qu’il a été démontré que des humains et des mammifères non humains décérébrés, c’est-à-dire à qui l’on a retiré la majeure partie du cerveau et en particulier le néocortex, continuent de présenter des comportements complexes qui vont au-delà du simple réflexe (5,6). Parmi ces comportements, on trouve l’apprentissage aversif et des comportements de protection visibles chez des rats décérébrés qui repoussent les seringues, tentent de les mordre et lèchent leurs blessures. Si ces comportements sont présents chez des individus décérébrés, donc probablement inconscients, ils ne peuvent pas être utilisés comme des preuves de douleur consciente lorsqu’ils sont observés chez des individus avec un cerveau intact. Pour la même raison, la diminution de ce type de comportement lors de l’administration d’analgésiques ne constituerait pas une preuve. En effet, l’analgésie fonctionne en diminuant la nociception précédant la douleur. Si les comportements évoqués peuvent être des réponses complexes mais inconscientes, leur diminution par analgésie ne constituerait pas une preuve de la présence de douleur.
Cependant cet argument part du principe que les individus décérébrés sont inconscients et donc ne ressentent pas la douleur, car ils ne disposent plus de néocortex. Ce second argument est donc sujet aux mêmes objections que le premier. Il est possible de le rejeter en supposant que ces animaux décérébrés ne sont peut-être pas totalement inconscients et que le néocortex, bien qu’incontestablement impliqué dans la conscience, ne serait pas la seule et unique structure responsable des expériences conscientes. Ainsi, certains ont avancé que le tronc cérébral, une structure apparue très tôt dans l’évolution des cerveaux et partagée par les humains et les poissons, pourrait aussi être impliqué dans la douleur (8). Cette hypothèse provient des cas d’enfants nés sans néocortex qui montrent tout de même des réactions à la douleur et des signes de conscience. Brian Key soutient en revanche que ces cas pourraient s’expliquer par la présence de restes parcellaires de néocortex chez ces enfants (6).
Notons toutefois que la position selon laquelle les poissons ne ressentiraient pas la douleur, voire ne seraient même pas conscients, est aujourd’hui minoritaire dans le monde académique. Ainsi, l’article de Brian Key de 2016 qui défendait cette thèse a généré pas moins de 30 articles de réponse, parmi lesquels 27 soutenaient que les poissons téléostéens ressentent probablement la douleur (7). De plus, des institutions scientifiques faisant autorité comme l’Institut national de recherche agronomique (INRA), l’Agence européenne de sécurité des aliments (EFSA), l’Organisation mondiale de la santé animale (OIE), l’Organisation mondiale de l’alimentation et de l’agriculture auprès des nations unies (FAO), et le Farm Animal Welfare Commitee (FAWC) – l’agence publique d’expertise sur le bien-être animal du Royaume-Uni –, considèrent que le niveau de preuve de la douleur des poissons est suffisamment élevé pour a minima leur accorder le bénéfice du doute et établir des recommandations visant à épargner des souffrances aux poissons, par exemple en les étourdissant avant abattage. Pareillement, le poisson zèbre, est couramment utilisé par de très nombreux chercheurs pour tester les médicaments psychiatriques destinés aux humains, ce qui suggère que le cerveau de ce poisson est jugé suffisamment proche du cerveau humain pour évaluer l’impact de substances influençant des états émotionnels similaires aux nôtres.
Une question éthique
Face au débat, Culum Brown, spécialiste du comportement des poissons, conclut de la manière suivante : « Nous devrions simplement accepter que nous ne pouvons pas ressentir ce que les autres animaux (ou les autres humains) ressentent, et que la question de la douleur chez les poissons revient à prendre des décisions prudentes et éthiques dans une situation d’incertitude : mettre en balance le poids des preuves en faveur et en défaveur de la présence de douleur chez les poissons, et le poids des conséquences d’un « faux négatif » consistant à considérer que les poissons ne ressentent pas la douleur s’ils la ressentent réellement ».
L’article 2 de la Déclaration des droits de l’animal avance que « Tout animal appartenant à une espèce dont la sensibilité est reconnue par la science a le droit au respect de cette sensibilité ». Dans le cas des poissons, de nombreuses pratiques devraient être modifiées pour atteindre cet objectif. Dans un article de 2011, Lynne Sneddon propose certaines pistes – reproduites dans le tableau suivant – pour aller vers une prise en compte de la sensibilité des poissons. Suivrons-nous ses éminents conseils ?
| Pratique | Sources de douleur | Pistes d’amélioration |
| Commerce des poissons d’ornements | _Blessures et stress lors de la capture en milieu naturel _Transport _Maladies et qualité d’eau médiocre | _Utilisation exclusive de poissons issus d’élevage _Réduction du temps de transport _Éducation des propriétaires |
| Pisciculture | _Densités importantes favorisant les comportements agressifs _Vaccinations à vif _Méthodes d’abattage | _Limitation des densités, choix des modes de nourrissage évitant la compétition entre les individus _Anesthésie avant vaccination _Étourdissement avant abattage |
| Pêche de loisir | _Blessures par les hameçons _Suffocation à l’air libre | _Utilisation d’hameçons sans crochet _Hameçons retirés sous l’eau avant de relâcher le poisson _Euthanasie des individus trop blessés pour être relâchés |
| Pêche commerciale | _Nombres importants d’individus soumis à la douleur _Utilisation de poissons en tant qu’appâts vivants _Suffocation à l’air libre ou éviscération lors de l’abattage | _Réduction du nombre de captures _Interdiction des appâts vivants _Étourdissement avant abattage |
| Expérimentation | _Procédures invasives (chirurgies, tests toxicologiques) | _Anesthésie et analgésie |
(Adapté à partir de Sneddon, 2011)
Conclusion
La douleur est une expérience sensorielle et émotionnelle désagréable. Comme il s’agit d’une expérience subjective « privée », il est impossible de savoir avec certitude si et comment elle est ressentie par autrui : humain ou animal. L’on peut toutefois observer le comportement, la physiologie et l’anatomie d’une espèce et se demander s’il paraît probable ou improbable que cette espèce ressente la douleur. Les preuves doivent être examinées à la lumière de nos connaissances sur la douleur humaine mais aussi en fonction de nos connaissances sur les processus de l’évolution des espèces. Bien qu’il existe des voix discordantes, nous sommes aujourd’hui proches du consensus sur l’existence de douleur chez les poissons téléostéens. Mais comme rien ne peut apporter de réponse définitive, il faut admettre qu’il s’agit avant tout d’une question éthique. Le poids des preuves doit être comparé aux conséquences morales de la prise en compte ou de la non prise en compte de la sensibilité potentielle d’une espèce.
Gautier Riberolles
2. Sneddon LU, Elwood RW, Adamo SA & Leach MC. (2014). Defining and assessing animal pain. Animal behaviour, 97, 201-212.
3. Sneddon LU. (2015). Pain in aquatic animals. Journal of Experimental Biology, 218(7), 967-976.
4. Sneddon LU. (2003). The evidence for pain in fish: the use of morphine as an analgesic. Applied Animal Behaviour Science, 83(2), 153-162
5. Rose JD, Arlinghaus R, Cooke SJ, Diggles BK, Sawynok W, Stevens ED, & Wynne CDL. (2014). Can fish really feel pain?. Fish and Fisheries, 15(1), 97-133
6. Key B. (2016). Why fish do not feel pain. Animal Sentience, 1(3), 1.
7. Brown C. (2016). Fish pain: An inconvenient truth. Animal Sentience: An Interdisciplinary Journal on Animal Feeling, 1(3), 32.
8. Brown C. (2016). Comparative evolutionary approach to pain perception in fishes. Animal Sentience: An Interdisciplinary Journal on Animal Feeling, 1(3), 5.


1 thought on “Douleur des poissons : va-t-on continuer à noyer… le poisson ?”
Les commentaires sont clos